
Why Longevity Interventions Fail Even When the Science Is Real
(Biological Readiness and the Missing Constraint Behind Durable Change)
By Brenden Henry

There are two kinds of skeptics.
One rejects anything that challenges the current paradigm, regardless of evidence.
The other is willing to ask uncomfortable questions and follow the data wherever it leads.
I’ve always been the second type.
My work in reversing aging didn’t start with supplements, protocols, or clinics. It started with regulatory peptides. I was studying molecules produced in nature that were designed to restore access to gene-expression programs that decline with age and disease. At face value, that idea sounds implausible to a lot of people. For a long time, I assumed it couldn’t be true.
But when I stepped outside conventional Western frameworks and focused on the biology itself, specifically on why systems lose the ability to regulate and repair, it became clear that these molecules did work. In some cases, they worked remarkably well.
And that’s where the real problem emerged.
Across different compounds, different mechanisms, and different people, the same thing kept happening.
The interventions would work, sometimes dramatically, but only under certain conditions. Even at identical doses, outcomes varied, and when they did work, the effects often didn’t hold.
That pattern led to a conclusion that signaling molecules are only as effective as the biological system prepared to receive them.
Most people who get serious about longevity eventually run into the same problem. An intervention works at first, a positive effect is noticed, sometimes even dramatically, and then it doesn’t last. The improvement plateaus, fades, or reverses, even though the intervention itself hasn’t changed. This happens with supplements, peptides, pharmaceuticals, and even stem cell therapies. The consistency of that pattern matters, because it tells us the failure is not intervention-specific. When fundamentally different tools break in the same way, the problem is almost never the tool. It’s the system receiving it.
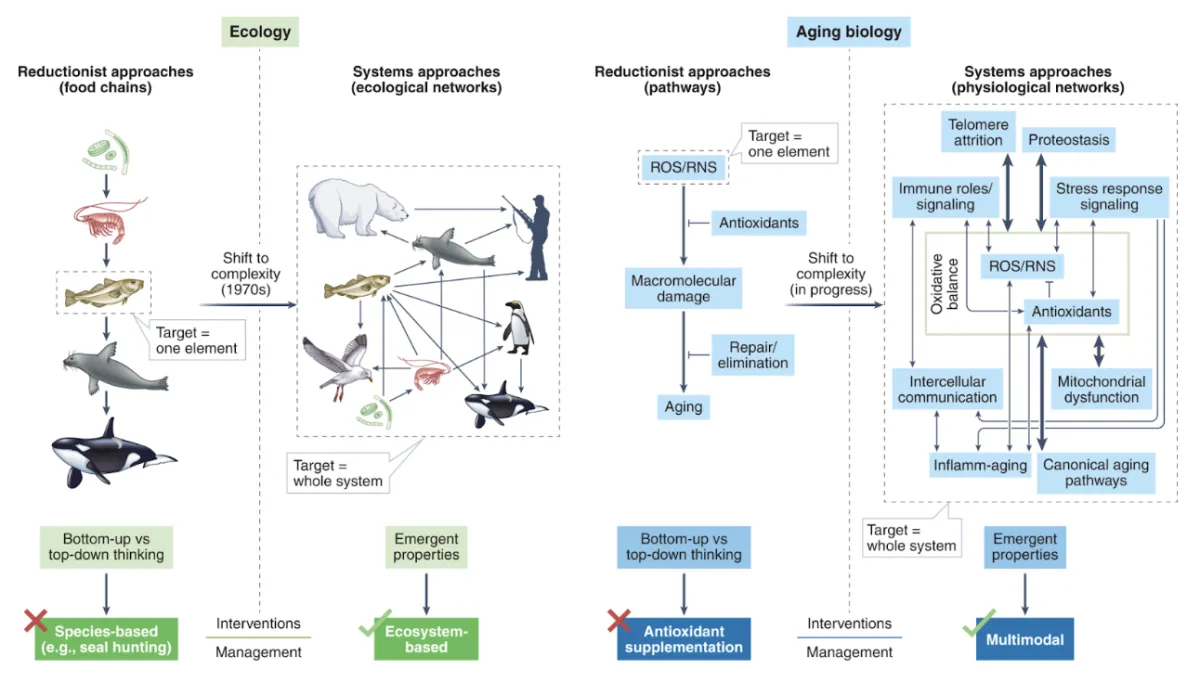
That constraint is not mysterious, but it is rarely discussed. For any biological intervention to produce a durable effect, the system has to meet three conditions simultaneously. It must have enough usable energy to respond without borrowing from the future. It must be able to interpret the signal clearly, without distortion or conflict. And it must be able to stabilize the resulting change so that it becomes a new baseline rather than a temporary deviation. If any one of those conditions fails, improvement is transient by definition.
This isn’t just a theory being put out there, its logic that is grounded in biology. Energy is not just calories or ATP — it’s mitochondrial efficiency, redox balance, immune cost, and metabolic reserve. When energy is marginal, the organism prioritizes survival and suppresses any attempts at adaptation in the way that you may hope with interventions. Signal clarity depends on intact receptors, DNA, feedback loops, and cross-system coordination between immune, endocrine, neural, and metabolic networks. When signaling is noisy, increasing the dose or layering additional interventions that aren't thought out carefully, doesn’t fix interpretation, it just amplifies distortion of the signal. Stability is governed by homeostatic set-points, including immune memory, tissue remodeling, and structural reserve capacity. Without stability, the body doesn’t lock the improvement in. It props it up temporarily which feels like success, until the intervention is reduced or removed.
This explains why escalation often feels necessary and why it also so often backfires. Escalation temporarily overcomes one constraint while worsening the others. Higher doses amplify signal but accelerate counter-regulation. Stronger interventions mobilize energy but deplete reserve. Regenerative inputs demand stability that isn’t present. The initial response is real, biology complies, but it does so by compensating, not by reorganizing. Compensation looks like progress until it collapses.
This is why the same intervention can feel powerful and then disappear. Biology is responding, but it is not reorganizing and consolidating those gains. Signals are being emitted, but the machinery required to interpret, amplify, and stabilize them is degraded. Without that machinery functioning properly, even correct inputs produce only transient effects.
It’s easy to assume the intervention itself will fix the machinery. In practice, that’s rarely what happens.
Stem cell therapies make this especially clear. They’re often treated as fundamentally different, as if living cells bypass the systemic limits of the organism receiving them. They don’t. Stem cells require functional circulation, immune signaling, metabolic energy, controlled inflammation, and a viable niche to integrate and persist.
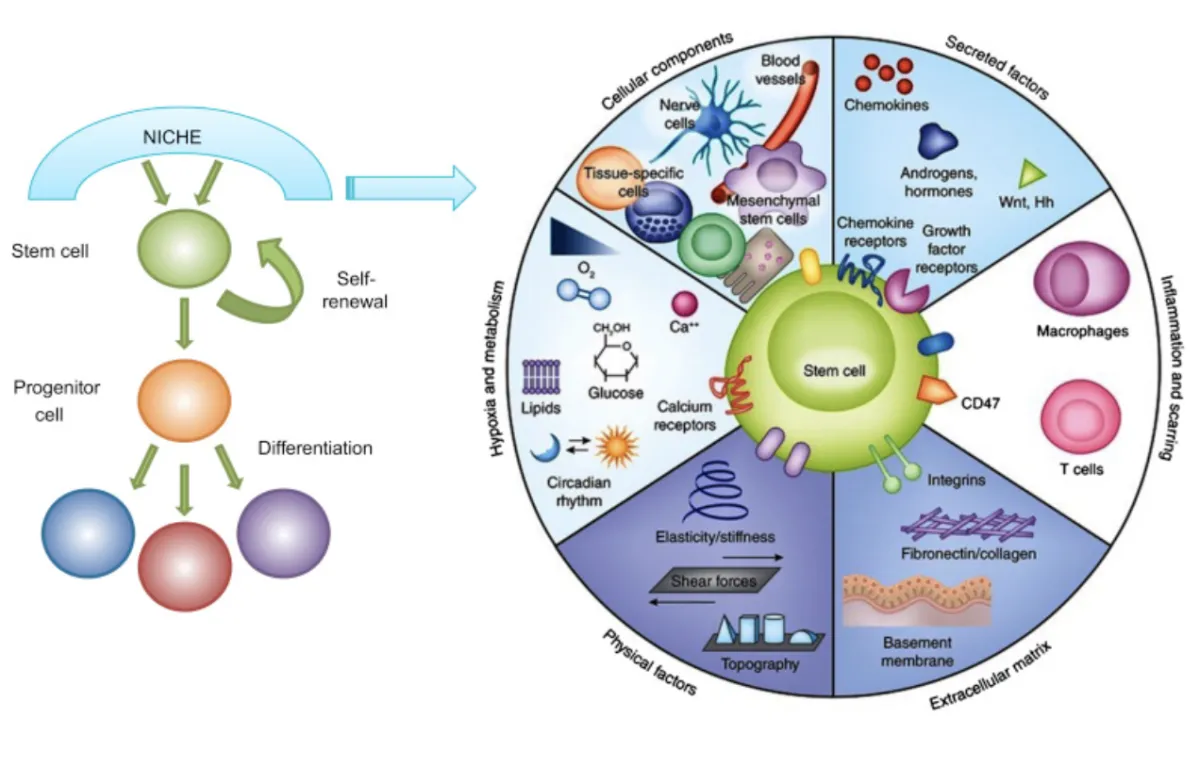
Without that, all you get is a transient burst of signaling the body can’t meaningfully use. When those conditions aren’t met, the cells do not persist for long. Think of it like planting expensive seeds into toxic soil.
Any benefit that follows is typically due to short-lived signaling, anti-inflammatory or trophic effects, not structural regeneration. That distinction matters, because signaling without integration cannot produce durable change. This outcome is not surprising, and it’s not a problem with the stem cells itself, but with the biology receiving the intervention without preparation. Autologous stem cells fail for the same reason, they’re introduced into the same environment that caused endogenous regeneration to decline and collapse in the first place. Nothing upstream changes.
What all of this points to is a missing variable that almost no one evaluates before intervening: biological readiness. Readiness determines whether a system can receive a signal, interpret it correctly, and hold the resulting change without destabilizing itself.
When readiness is low, the pattern is always the same. Interventions do something, but they don’t settle. Gains require constant reinforcement. The system becomes dependent on timing, dosing, and escalation just to maintain baseline function. Miss a step, and the effect disappears. That isn’t optimization, it’s fragility.
When readiness is restored, the behavior of the system changes. Interventions no longer need to overpower resistance. Smaller inputs produce clearer effects. Improvements persist even when the intervention is reduced or removed. The system starts to cooperate instead of push back.
This is where most modern longevity efforts go wrong. They focus almost entirely on optimization, improving markers, activating pathways, increasing output, without asking whether the system can actually support those changes long term. The result is improvement that only holds as long as the intervention is constantly applied. Optimization increases demand on the system. Readiness determines whether that demand can be met.
This is not an argument against optimization. Optimization is necessary. It’s just downstream.
When governance comes first and readiness is restored, optimization stops being fragile. Markers improve without constant escalation. Pathways respond without being forced. Output increases without depleting reserve.
In a governed system, optimization finally behaves the way the science says it should.
That’s what governance actually means. It’s the difference between managing interactions and simply adding force. Governance means sequencing pressure appropriately and protecting future capacity instead of consuming it. Optimized systems require constant intervention just to hold gains. Governed systems don’t. They stabilize improvements, tolerate variation, and stop treating every missed input as a crisis.
When biology is governed, responses become predictable. Improvements stabilize instead of oscillating. Escalation becomes unnecessary because the system is no longer resisting or counter-regulating the intervention.
The widespread failure of longevity interventions isn’t evidence that the science is fake or incomplete. It’s evidence that downstream tools are being applied to systems that no longer have the structural permission to change. Until those upstream constraints are addressed, improvement cannot hold no matter how advanced the intervention appears.
That’s why The Biological Recode System exists. It’s designed to identify and restore biological readiness and to optimize biology at the system level by governing how pathways are activated, how interventions are combined, and how pressure is applied over time.
When biology is governed, optimization stops being so fragile. Instead it becomes stable, compounding, and predictable. Markers improve without constant escalation. Pathways respond without being forced. Output increases without depleting reserve.
That’s the difference between people who are chasing effects and achieving real biological optimization.
After years of watching this exact pattern repeat, in myself, my family, and across hundreds of clients, it became clear that adding more interventions wasn’t the answer. The problem wasn’t a lack of tools. It was the absence of governance.
That’s why I built the Biological Recode System: to govern biology, assure readiness, and deliver optimization at a level no standalone interventions can achieve.
If you want to apply for access to the Biological Recode System, there’s a link below:
To Rewriting Your Biology,
Brenden Henry
Former Biomedical Engineer
Founder of Peptide Science Institute &
CuttingEdgeLongevity

support:
sms +1 877-834-9117
email [email protected]